Как называется перетяжка соединяющая хроматиды
Слово «хромосома» означает «окрашенное тело» (окрашивается более эффективно, чем остальные компоненты клетки). Каждая хромосома состоит из очень длинной молекулы ДНК, связанной с РНК и белками, образующей вещество, известное как хроматин. Во время интерфазы клеточного цикла (когда клетка не делится) хромосомы распределены по всему ядру, в то время как при митозе и мейозе они уплотняются, располагаясь ближе друг к другу.
ДНК содержится именно в хромосомах, вероятно, потому, что это облегчает разделение полного набора генов между дочерними клетками на стадии митоза, а также её упаковку в головки сперматозоидов после мейоза.
Способность к окрашиванию используют в диагностических целях для визуального исследования хромосом, их идентификации и выявления патологий. Различают слабоокрашенные части (эухроматин) и области с более выраженной окраской (гетерохроматин).
Генетическая информация (геном) заключена в молекуле ДНК и кодирована определённой последовательностью азотистых оснований в ДНК. Основная часть этой информации находится в составе хромосом в ядре клетки, однако небольшое количество ДНК в виде свободных цепочек локализовано в митохондриях цитоплазмы. Ядра присутствуют практически во всех клетках организма человека, за исключением эритроцитов и клеток хрусталика.
Обычно в каждом ядре содержится двухметровая цепь ДНК, разделённая между 23 парами хромосом (примерно по 4 см на хромосому). Однако непосредственно перед делением она уплотняется до 5 um (0,005 мм) посредством сложных процессов спирализации и уплотнения.
Структура хроматина хромосом
В каждой хромосоме цепь ДНК дважды обмотана вокруг октамерного комплекса, состоящего из восьми «белков хранения» (гистонов), формирующих нуклеосомы, при этом образованная структура напоминает «бусы на нити». Кора (сердцевина) нуклеосомы состоит из двух молекул каждого из четырех гистонов— Н2А, Н2В, НЗ и Н4. Гистоны заряжены положительно, а потому могут образовывать ионные связи с отрицательно заряженными фосфатными группами ДНК.
Последовательность аминокислот в гистонах у всех видов совпадает практически на 100%, указывая на огромную значимость данных элементов в поддержании структуры и функций хроматина. Каждая нуклеосома вмещает порядка 200 пар оснований молекулы ДНК и укорачивает длину цепи ДНК на одну десятую.
Затем подобная бусам нить спирализуется в соленоид, или зигзагообразную спираль (фибрилла длиной 30 нм), состоящую из 5—6 нуклеосом. При этом её структура поддерживается одной молекулой гистона HI на каждую нуклео-сому. Коэффициент упаковки при образовании соленоида примерно равен 5, что в свою очередь даёт общий коэффициент упаковки 50. Существует предположение, что в таком состоянии эухроматин находится во время интерфазы в местах, где гены не экспрессируются.
Во время митоза и мейоза хромосомы ещё больше уплотняются, при этом коэффициент упаковки по отношению к предыдущим уровням составляет 100, а общий коэффициент может достигать 5000. Считают, что фибриллы хроматина складываются в вытянутые петли, в основании которых расположен скаффолд (остов) из негистоновых хромосомных белков, которые прикрепляются к определённым последовательностям оснований, рассредоточенным вдоль молекулы ДНК. Сжатие этих негистоновых хромосомных белков предположительно обусловливает уплотнение хромосом.
Один из ключевых белков скаффолда — топоизомераза II, фермент, который расщепляет двойные цепи ДНК, переносит их через разрыв и вновь соединяет, что в свою очередь необходимо для релаксации суперспиралей ДНК во время репликации или транскрипции. Топоизомераза II прикрепляется к участкам связывания со скаффолдом, которые богаты А- и Т-основаниями (более 65% оснований составляют А и Т, см. главу 4). Считают, что каждая петля выступает в роли независимого функционального домена во время репликации или транскрипции ДНК.
На следующем этапе петли фибрилл хроматина спирализуются, в результате чего возникает наиболее уплотнённый гетерохроматин, входящий в состав хромосомы во время деления клеток.
Дифференциальное окрашивание хромосом
Наиболее уплотнённые части хромосом хорошо окрашиваются красителем Гимзы (G-окраска). При этом отчётливо видны сильно уплотнённые маленькие петли (G-полосы), так как участки связывания со скаффолдом тесно прилегают друг к другу. Их репликация происходит во время S-периода, они не активны во время транскрипции. Полосы, которые плохо окрашиваются раствором Гимзы, или R-полосы, состоят из более свободных петель, богатых Г- и Ц-основаниями, они отличаются высокой активностью во время транскрипции. В основе идентификации хромосом лежат различия в их окраске.
Центромера хромосом
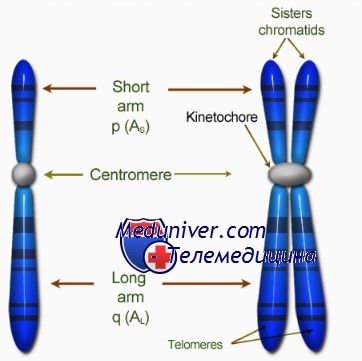
Видимые на ранних стадиях митоза хромосомы состоят из двух абсолютно одинаковых структур, называемых сестринскими хроматидами, которые соединяет первичная перетяжка. Она представляет собой недуплицированный участок ДНК, или центромеру, которая дуплицируется в начале анафазы митоза.
Кинетохор — органелла, которая локализована на каждой стороне каждой центромеры в период ранней профазы митоза и облегчает полимеризацию димеров тубулина, необходимую для образования микротрубочек митотического веретена (веретена деления).
Тепомера хромосом
Теломерой называют специализированный участок конца хромосомы. К ней прикрепляются теломерспецифические белки, образующие «шапочку» (cap) для защиты конца хромосомы.
Теломеры предположительно препятствуют патологическому слиянию концов хромосом конец в конец, обеспечивают полноту репликации, участвуют в образовании пар хромосом во время мейоза и помогают восстанавливать внутреннюю структуру ядра на стадии интерфазы путём присоединения хромосом к ядерной мембране.
Эухроматин и гетерохроматин хромосом
Эухроматин находится в уплотнённом состоянии во время деления клеток и распаковывается на стадии интерфазы. Он обусловливает бледное окрашивание R-полос в уплотнённых хромосомах и содержит большинство структурных генов.
Гетерохроматин очень сильно уплотнён во время деления клетки и остаётся в таком состоянии даже на стадии интерфазы. Он локализован в основном на периферии ядра и около ядрышка и не активен во время транскрипции. Конструктивный (структурный) гетерохроматин одинаков во всех клетках организма, в то время как факультативный гетерохроматин различен в зависимости от типа генов, экспрессируемых в дифференцированных клетках определённых тканей.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
На пути к разгадке тайны мейоза

Появление мейоза – особого варианта клеточного деления, в результате которого число хромосом сокращается вдвое – было одним из важнейших эволюционных «достижений» первых эукариот. Механизм мейоза сложился из комбинации готовых «блоков»: механизмов митоза, рекомбинации и репарации ДНК. Ключевым событием стало формирование синаптонемного комплекса – особой белковой структуры, обеспечивающей попарное соединение и точное «выравнивание» хромосом.
Появление эукариотической клетки было важнейшим эволюционным преобразованием (ароморфозом) в истории земной жизни (см. обзор «Происхождение эукариот» ). Одним из главных «достижений» древних эукариотических организмов стало возникновение настоящего полового процесса, то есть слияния двух гаплоидных (содержащих одинарный набор хромосом) клеток – гамет в диплоидную (содержащую двойной набор хромосом) клетку – зиготу.
Чтобы жизненный цикл эукариот, обладающих половым процессом, мог продолжаться, должен был развиться механизм, посредством которого из диплоидных клеток снова могли образовываться гаплоидные. Таким механизмом стал мейоз – особый вид клеточного деления, при котором число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой.
В статье Ю.Ф.Богданова, крупнейшего специалиста по эволюции мейоза, рассматриваются современные представления о происхождении механизмов мейотического деления у эукариот.
Общепризнано, что мейоз произошел из митоза – «обычного» клеточного деления эукариот, в результате которого число хромосом остается прежним. Происхождение митоза само по себе было замечательным ароморфозом. Мейоз, по сути дела, является модифицированной версией митоза.

В ходе митоза (а также в ходе второго деления мейоза) хромосомы вступают в метафазу поодиночке, и нити веретена деления присоединяются к каждой хромосоме с двух сторон.
Именно благодаря указанным особенностям первого деления мейоза и обеспечивается уменьшение числа хромосом: к полюсам клетки расходятся не сестринские хроматиды, а гомологичные хромосомы, по одной из каждой пары (см. схему).
Эти ключевые особенности первого деления мейоза развились на основе одного и того же ароморфоза – возникновения так называемого синаптонемного комплекса (СК).
Механизм мейоза был «собран» из готовых блоков (как это часто бывает в эволюции) – разумеется, с некоторыми модификациями и новшествами.
Одним из этих исходных блоков стал уже имевшийся к тому времени у эукариот механизм митоза, другим – механизм репарации (починки повреждений) ДНК, основанный на гомологичной рекомбинации (см.: Глазер В.М. Гомологичная генетическая рекомбинация ). Суть процесса в том, что поврежденный участок одной молекулы ДНК заменяется его неповрежденной копией, взятой из другой (гомологичной) молекулы ДНК.
Чтобы «развести» гомологичные хромосомы к разным полюсам клетки, в ходе мейоза используется веретено деления, «унаследованное» от митоза. Но чтобы распределение хромосом по дочерним клеткам прошло без ошибок, гомологичные хромосомы необходимо сначала сгруппировать попарно. Для этого используется механизм гомологичной рекомбинации, точнее, одна из деталей этого механизма – образование так называемого соединения Холлидея.

Починка разрывов ДНК – одна из основ мейоза
Механизмы репарации и рекомбинации ДНК – очень древние, они должны были возникнуть еще на заре жизни.
Становление мейоза в ходе эволюции, так же как и сам мейоз в жизненном цикле современных организмов, начинается с репарации повреждений (разрывов) ДНК. Разрывы эти в начале мейоза возникают не сами собой – их создает специальный фермент, эндонуклеаза SPO11. Этот фермент является модификацией другого белка – ДНК-топоизомеразы VI, унаследованной эукариотами от своих прокариотических предков – архей.
Клетка начинает «чинить» разорванные молекулы ДНК при помощи древнего механизма гомологичной рекомбинации. А для этого нужно объединить попарно гомологичные молекулы ДНК (чтобы использовать неповрежденные участки одной молекулы в качестве «матрицы» для исправления повреждений в другой молекуле). На этом и основано попарное объединение хромосом в профазе I мейоза.
Белковые оси хромосом
Эукариотический геном значительно больше прокариотического и содержит больше «избыточной» ДНК (различных некодирующих последовательностей, мобильных генетических элементов и др.) Большие размеры генома стимулировали «распадение» его на отдельные хромосомы (линейные, в отличие от единственной кольцевой хромосомы прокариот). Как следствие, «оказалась полезной новация в структуре хромосом – белковые оси, к которым стали крепиться петли гигантской молекулы ДНК – эукариотической хромосомы».
В клетках эукариот, делящихся путем митоза, сестринские хроматиды (образовавшиеся в результате репликации исходной молекулы ДНК) соединены посредством белков-когезинов. Чтобы хроматиды могли разойтись к полюсам клетки (в анафазе митоза), когезины должны гидролизоваться и исчезнуть из межхроматидного пространства.
Один из когезинов (Rec8) «оказался подходящим субстратом для наслоения на него других мейоз-специфичных белков», из которых в начале мейоза строятся более сложные белковые оси, или «линейные элементы», к которым ДНК крепится петлями, приобретая вид «ершика». Каждая петля содержит несколько тысяч пар нуклеотидов и крепится к линейному элементу только в одном месте, «кнопочно». Такая организация хромосомы позволяет хромосомным локусам легче находить гомологичные им локусы в хромосоме-партнере. Так обстоит дело у некоторых низших эукариот, у которых нет «классического» мейоза, характерного для более высокоорганизованных форм.
Синаптонемные комплексы. Ароморфоз.
У эукариот с классическим мейозом «бывшие линейные элементы хромосом соединяются попарно с помощью белковой «застежки-молнии» в двухосевую структуру, именуемую синаптонемным комплексом (СК)». Это обеспечивает тесное соединение гомологичных хромосом. Ю.Ф.Богданов считает появление СК важным ароморфозом. СК существует ограниченное время в течение профазы I мейоза и затем распадается. СК обнаружен у многих сотен видов – от одноклеточных водорослей, грибов и протистов до многоклеточных. Предполагается несколько возможных функций СК:
1) Организация профазной мейотической хромосомы. Благодаря СК возникает билатеральная организация пары соединившихся гомологичных хромосом. Петли хроматина располагаются по обе стороны от СК. Это позволяет точно сопоставить петли хроматина в трехмерном пространстве клеточного ядра. «По-видимому, это простейший, если не единственный, способ внести порядок и точность в процесс взаимного узнавания локусов гомологичных хромосом и обеспечить достаточную точность (гомологию) рекомбинации в мейозе.»
2) СК не только соединяет гомологичные хромосомы, но и не дает им «склеиться», удерживая их на расстоянии 70-120 нм друг от друга. В конце профазы I (стадия диплотены) СК распадается во всех локусах, кроме локусов хиазм (перекрещивания хроматид). Гомологичные хромосомы, взаимно оттолкнувшиеся во всех локусах, кроме локусов хиазм, выстраиваются на экваторе веретена деления в метафазе I. Они готовы разойтись к полюсам, как только освободятся от хиазм.
3) СК необходим для формирования хиазм. СК также не позволяет хиазмам располагаться слишком близко друг от друга. Иными словами, благодаря СК хроматиды могут «рваться» и обмениваться участками лишь в ограниченном количестве мест. Ю.Ф.Богданов оценивает это как положительное явление, поскольку «чрезмерная частота кроссинговера – не столько «благо» комбинаторики, сколько угроза стабильности адаптационно выгодных фенотипов».
Происхождение белков СК
У разных групп эукариот СК строится по единому «плану», но при этом используются совершенно разные (структурно негомологичные) белки. Это похоже на постройку домов по сходному плану (стены, крыша, окна), но из совершенно разных материалов. Материал не важен – «важно, чтобы СК выравнивал параллельно лежащие гомологичные хромосомы, сохранял между ними пространство, в котором происходит рекомбинация ДНК, и сохранял бы эти условия столь долго, сколько необходимо для завершения рекомбинации и формирования хиазм».


Сравнительный анализ структуры белков СК показал, что они возникли независимо у предков современных эукариот, давших начало разным крупным таксонам (растений, грибов, животных).
Ю.Ф.Богданов приводит убедительные доводы и экспериментальные факты, свидетельствующие о том, что структуры СК формируются путем самосборки из белковых молекул, подобно многим другим сложным молекулярным комплексам. Автор отмечает, что «самоорганизующиеся структуры могут состоять из разных белковых субъединиц и нуклеиновых кислот, как это наблюдается в отношении вирусов и рибосом». Важно, что белки, сходные по размерам и трехмерной организации, могут служить компонентами одинаковых структур у далеких друг от друга организмов даже в тех случаях, когда первичная структура (аминокислотная последовательность) этих белков сильно различается.
Почему в первом делении мейоза сестринские хроматиды не расходятся?
Итак, причина нерасхождения хроматид в мейозе I – сохранение соединяющей их когезиновой оси. В результате после первого деления мейоза «программа» клеточного деления остается как бы «не выполненной». Эта программа – в основе которой лежит древняя программа митоза – после завершения мейоза I, не останавливаясь, продолжает работать (хроматиды не разошлись, когезиновые оси не гидролизовались – значит, нужно делиться дальше). И поэтому клетки сразу же, без интерфазы и без синтеза ДНК, вступают в новое деление (мейоз II), идущее уже в точности по схеме классического митоза.
В заключительнй части статьи Ю.Ф.Богданов рассматривает ряд интересных случаев отхода некоторых организмов от схемы «классического» мейоза. Например, у самцов дрозофил (в отличие от самок) СК не образуется, и мейоз идет по примитивной схеме, характерной для некоторых архаичных эукариот (дрожжи S. pombe ). Таким образом «генетическая программа» примитивного мейоза у высших организмов не исчезает (вероятно, она просто входит в состав более сложной программы «классического» мейоза) и может вновь оказаться востребованной.
Ю. Ф. Богданов. Белковые механизмы мейоза (популярная статья в журнале «Природа»).
С фактами и теориями, касающимися происхождения мейоза, можно ознакомиться в статьях Ю.Ф.Богданова и его коллег:
Что такое ДНК и хромосомы
Что такое ДНК, и из чего она состоит? Кто и когда открыл эту молекулу в клетках человека и других живых организмов? Чем уникален открытый учеными механизм наследования, и какие последствия ждал весь мир после этого открытия? Всю необходимую информацию Вы можете узнать, прочитав эту статью.

Когда впервые в истории появилось упоминание о ДНК
Иоганнес Фридрих Фишер – врач и биолог-исследователь родом из Швейцарии, стал первым в мире ученым, выделившим нуклеиновую кислоту. Открытие случилось в 1869 году, когда он занимался изучением животных клеток, а именно лейкоцитов, которых много содержалось в гное. Совершенно случайно молодой ученый заметил, что при отмывании лейкоцитов с гнойных повязок от них остается загадочное соединение. Под микроскопом Иоганн обнаружил, что оно содержится в ядрах клеток. Это соединение Мишер назвал нуклеином, а в процессе изучения его свойств переименовал в нуклеиновую кислоту, из-за наличия свойств, как у кислот.
Роль и функции только открытой нуклеиновой кислоты были неизвестны. Однако многие ученые того времени уже высказывали свои теории и предположения о существовании механизмов наследования.
Нынешние взгляды на состав молекулы ДНК ассоциируются у людей с именами английских ученых Джорджа Уотсона и Фрэнсиса Крика, которые открыли структуру данной молекулы в 1953 году. За несколько лет до этого, в тридцатые годы, ученые из советского союза А.Н. Белозерский и А.Р. Кезеля доказали наличие ДНК в клетках во всех живых организмах, тем самым они опровергли теорию о том, что молекула ДНК находится только в клетках животных, а в клетках растений присутствует только РНК. Лишь спустя несколько лет, в 1944 году, группой освальдских ученых было установлено, что молекула ДНК является механизмом сохранения наследственной информации клетки. Таким образом, благодаря совместным усилиям и трудам исследователей человечество познало тайну процесса эволюции и его основных принципов.
ДНК в медицине
Открытие состава молекулы дезоксирибонуклеиновой кислоты позволило перейти медицине на новый уровень развития. Появилось большое количество новых направлений практической медицины, стали доступны новые методы лечения, диагностики. Благодаря этому фундаментальному открытию для науки и современным технологиям, человечеству стали доступны:
И это еще не все доступные для людей услуги, которые может предложить медицина, изучающая генетику. Выше были представлены только самые популярные среди людей тесты. Перспективой для многих ученых-генетиков является создание таких лекарств, способных победить все болезни на Земле и даже смертность.
Строение молекулы ДНК
От цепочки к хромосоме
В каждом живом организме находится миллионы клеток, а внутри этих клеток находится ядро. Клетки, содержащие в себе ядро, называются эукариотами или ядерными. У древних одноклеточных нет оформленного ядра. К таким безъядерным одноклеточным, или прокариотам, относятся бактерии и археи, например, кишечная палочка или серая анаэробная бактерия. Также ядро отсутствует в клетках вирусов и вироидов, однако причисление вирусов к живым организмам – вопрос спорный, о котором по сей день дискуссируют ученые.
В ядре находятся хромосомы – структурный элемент, в котором содержится молекула ДНК в виде спирали, хранящая внутри себя всю генетическую информацию клетки.
Процесс упаковки ДНК спиралей
Количество нуклеотидов в ДНК велико, и нужны длинные цепочки, чтобы вместить все их число, поэтому нити ДНК закручиваются в две спирали, что позволяет укоротить цепочки в 5 раз, сделав их более компактными. Нити ДНК могут также закручиваться в форму суперспирали. Двойная спираль пересекает свою ось и накручивается на специальные гистоновые белки – гиразы, образуя при этом супервитки. Таким образом, двойная спираль закручивается в спираль более высокого порядка. Сокращение цепочек в этом случае произойдет в 30 раз.
Как гены связаны с ДНК
Ген – самый изученный на сегодняшний день участок ДНК. Гены являются структурной единицей наследственности всех живых организмов. Цепочки нуклеотидов в ДНК состоят из генов, которые определяют генотип особи, например, цвет и разрез глаз, тип кожи, рост, группу и резус фактор крови и другие физиологические качества и особенности внешности.
Еще много отраслей генетики до конца не изучены, и до конца не раскрыты все функции генома, но ученые до сих пор продолжают изучение генов, чтобы добиться новых открытий в области генетики.
Хромосома: определение и описание

Хромосомы – структурный элемент клетки, находящийся внутри ядра. Они содержат в себе молекулы ДНК, в которых содержится вся наследственная информация.
Строение и виды хромосом:
Отсюда возникают различные типы хромосом:
Всего в клетке человека находится 46 хромосом: 22 пары аутосом, встречающиеся у обоих полов, и одна пара половых хромосом: XY – у мужчин, XX – у женщин. Забавно, что если прибавить к количеству хромосом хотя бы одну пару, то человек мог бы быть шимпанзе или тараканом, а если отнять, то – кроликом.
Еще интересно то, что человек и ясень имеют одинаковое количество хромосом, несмотря на принадлежность к разным видам и царствам.
Наследственные болезни
Генетический код – система записи генетической информации в ДНК и РНК в виде определенной последовательности в цепочке нуклеотидов. Он должен сохранять наследственную информацию в первоначальном виде, восстанавливая повреждения цепочки в последующем поколении с помощью ДНК. Однако ген может каким-то образом быть поврежден, либо в нем может произойти мутация.
Генные мутации – изменение в последовательности нуклеотидов, например выпадение, замена, вставка другого нуклеотида в цепочку. Последствия этих мутаций могут быть полезные, вредные или нейтральные. Примером полезных мутаций является устойчивость к минусовым температурам, увеличенная плотность костей, меньшая потребность во сне, устойчивость к ВИЧ и другие. Примером вредных мутаций является аллергия на солнечный свет, глухота слепота и так далее. К нейтральным мутациям относятся те мутации, которые не влияют на жизнеспособность, например, гетерохромия.
Существуют также летальные и полулетальные мутации. Летальные мутации несовместимы с жизнью и приводят к гибели организма на ранних этапах его развития, например, при рождении у особи отсутствует головной мозг. Полулетальные мутации не приводят к смерти особи, но значительно уменьшают ее жизнеспособность. К таким мутациям относятся заболевания человека, передающиеся по наследству. Например, наличие 47-й хромосомы может вызвать у человека синдром Дауна, а, наоборот, отсутствие 46-й парной хромосомы – сидром Шерешевского-Тернера.
Расшифровка цепочки ДНК
Расшифровка цепочки ДНК в клетке – это исследование всех известных генов в клетках человека. Хоть цена за такую услугу значительно упала за последние десять лет, однако такое исследование по-прежнему остается дорогим удовольствием, и не каждый человек сможет позволить себе оплатить такую услугу. Чтобы уменьшить цену этого исследования, расшифровку ДНК стали делить по тематикам. Таким образом, появились различные тесты, которые исследуют интересующую человека группу генов и ее функции.
Как происходит расшифровка цепочки ДНК?
Таким образом, ученые получают картину гена, которую можно изучить и расшифровать. Синтез РНК Нуклеотиды делятся на четыре базовых элемента, служащими основой для формирования генов: АТГЦ, или аденин, тимин, гуанин, цитозин. В их состав входят фосфорные остатки, азотистые основания и пептоза.
Важно, что молекула дезоксирибонуклеиновой кислоты не должна выходить за пределы мембраны ядра. С помощью РНК, которая играет роль копии участка цепи с генетическим кодом, генетическая цепочка может покинуть ядро, попасть вовнутрь клетки и воздействовать на ее внутренние процессы.
Как это происходит:
Итак, группа генов, участвующих в процессе старения клеток может, как заставить процесс старения идти быстрее, так и вовсе его остановить и запустить процесс омолаживания. То есть, каждый из генов может спровоцировать синтез нескольких видов белка.

Сутягина Дарья Сергеевна
В нашей ДНК содержится очень много информации, но пока мы можем расшифровать лишь небольшой процент генов. Добавлю несколько интересных фактов о ДНК: возможность двойной ДНК у человека. Такое явление случается, когда при беременности в утробе развиваются близнецы, но в процессе развития плода они сливаются в одного человека. Длина одной молекулы ДНК человека равна 2 метрам, а общая длина цепочки ДНК всех клеток тела человека равна 16 млрд. километрам, что равно расстоянию от Земли до Плутона. ДНК человека и кенгуру всего лишь 150 млн. лет назад были одинаковыми. Все знания и информация во всем мире могла бы уместиться всего лишь в 2 граммах дезоксирибонуклеиновой кислоты.
ООО «Медикал Геномикс» Лицензия № ЛО-69-01-002086 от 06.10.2017
Юр. адрес: г. Тверь, ул. Желябова, 48
ООО «Лаб-Трейдинг», ИНН: 6950225035, ОГРН: 1186952017053, КПП:695001001
Юр. адрес: г. Тверь, ул. 1-Я За Линией Октябрьской Ж/Д, 2, оф. 22



